
Extrait de Vivre parmi les animaux, mieux les comprendre de Pierre Le Neindre et Bertrand L. Deputte
Il est possible d’analyser le contenu de la conscience. Pendant très longtemps, il a été admis que seuls les humains possédaient un contenu de conscience. Celui-ci était d’ailleurs associé à une compétence morale, aux sens de bien et de mal. Cette compétence morale a été mise de côté dans ce texte, car sans pertinence pour les animaux.
Les techniques et concepts développés au cours des 20 dernières années remettent en cause le dogme selon lequel seuls les humains ont une conscience telle qu’elle est définie, c’est-à-dire une réflexivité de l’individu sur lui-même et sur ce qu’il fait. Différents travaux expérimentaux ont permis d’explorer sur certaines espèces plusieurs expressions de cette conscience. Dans le cadre de l’expertise sur la conscience, différentes expressions ont été analysées. Elles seront rappelées brièvement ici, tels les émotions, la métacognition, le voyage dans le temps et les relations entre les individus.
Au-delà de la réponse simple des animaux à des stimuli, il est important de mettre en avant les émotions des animaux en réponse à un évènement et sa valence, c’est-à- dire l’intérêt que l’animal lui porte, négativement ou positivement. Ces émotions supposent une analyse de la situation par rapport à des attentes qui ne nécessite pas forcément des processus cognitifs complexes (Boissy et al., 2007). Des études expérimentales conduites en particulier sur des ovins ont montré l’importance d’une composante subjective qui détermine des réponses comportementales et physiologiques spécifiques. Différentes modalités, dont la pertinence et les implications pour l’individu, les possibilités d’action et la signification normative, modulent les réactions du sujet. Il a été montré que les animaux tiennent compte de leurs expériences passées et se projettent dans l’avenir. Ils ressentent donc des états émotionnels comparables à ceux étudiés chez les humains.
Des expériences positives ou négatives modifient l’apprentissage et la mémorisation des ovins. En particulier l’accumulation d’expériences émotionnelles négatives ou positives module l’évaluation des situations auxquelles les animaux sont soumis ultérieurement. Ceci les conduit par exemple à avoir des attitudes « optimistes » ou « pessimistes », ou biais cognitif, lors de leur confrontation avec des situations nouvelles (Destrez et al., 2013 et 2014). Ce biais cognitif a également été démontré chez les porcs (Douglas et al., 2012).
Un autre exemple intéressant à considérer est celui de la mise en évidence d’une mémoire épisodique chez les animaux. Cette capacité a pendant longtemps été considérée comme étant le privilège de l’homme. La mémoire épisodique décrit le fait que nous pouvons nous souvenir d’évènements particuliers de notre vie. Elle se distingue de la mémoire sémantique qui porte sur la connaissance des règles, du style « le matin je me lève » ou « le soleil se lève à l’est le matin ». Cette mémoire épisodique a été tout particulièrement étudiée chez les humains. Certains humains peuvent d’ailleurs avoir une mémoire sémantique sans avoir de mémoire épisodique. On sait par ailleurs qu’une structure nerveuse, l’hippocampe, est impliquée dans cette mémoire.
L’absence de preuve de mémoire épisodique chez les animaux a conduit certains auteurs à conclure qu’ils étaient « coincés dans le présent » (Roberts, 2002, stuck in time). Cette position était assez largement partagée par les philosophes et scientifiques jusqu’à une période récente. Les maîtrises du passé et de l’avenir étaient données comme une des spécificités d’humains adultes qui seuls pouvaient exprimer leurs souvenirs et prévisions. La première brèche dans ce dogme a été de constater que cette proposition n’était pas falsifiable pour les animaux puisqu’il fallait qu’ils explicitent leurs souvenirs alors qu’ils n’ont pas de langage. Il a fallu définir des méthodes qui donnaient des résultats falsifiables sans le langage. Pour rendre les hypothèses falsifiables pour les animaux, les chercheurs du domaine ont convenu qu’un évènement pouvait être caractérisé par trois composantes : quoi, où et quand, ou « QOQ » (en anglais WWW pour what, where, when).
Une des premières démonstrations de l’opérationnalité de cette démarche a résulté d’expérimentations sur un corvidé nord-américain, le geai buissonnier de l’Ouest (Aphelocoma californica). Cet oiseau vit dans un milieu où les ressources sont rares et saisonnières. Une deuxième caractéristique de ces geais est le comportement de cache des aliments. Un dispositif simple, mais ingénieux, a permis de tester les hypothèses des auteurs. Dans le contexte expérimental, ces oiseaux préfèrent consommer des larves d’insectes, qui se dégradent rapidement, plutôt que des granulés, qui ne se dégradent pas dans le temps. Les animaux qui étaient habitués à vivre dans des cages avaient la possibilité de cacher dans des bacs soit des larves d’insectes, soit des pellets. Après les avoir laissés cacher des aliments des deux types dans des bacs remplis de sable, les oiseaux ont été retirés de la salle. Ils y ont été replacés soit quelques heures après, soit des jours après. Dans le deuxième cas, les larves étaient mortes et immangeables. Après un délai court, ils ont cherché préférentiellement là où ils avaient caché des larves. En revanche, après un délai long, ils ont cherché dans les endroits où ils avaient cachés des pellets. Ils démontraient ainsi leurs capacités à répondre au paradigme QOQ (Clayton et Dickinson, 1998). Bien que cette conclusion ait été combattue avec vigueur par certains en particulier (Süddendorf et Busby, 2003), l’accumulation de données pour répondre aux critiques a conduit à accepter cette conclusion. Les échanges entre les scientifiques sont rapportés dans l’expertise collective sur la conscience des animaux (Le Neindre et al., 2017).
Des observations en nature et des expérimentations au laboratoire ont permis de conforter ces conclusions. D’autres espèces, comme les écureuils, de nombreux rongeurs et certaines mésanges, ont la même habitude de cacher des aliments. Des dispositifs semblables ont été par la suite mis au point sur différentes espèces, dont les rats (Eacott et al., 2005), les porcs (Kouwenberg et al., 2009), les colibris ou Selasphorus rufus (Jelbert et al., 2014), la pie bavarde (Zinkivskay et al., 2009) et les campagnols des prairies ou Microtus pennsylvanicus (Ferkin et al., 2008). Pour cela, les auteurs ont pris en compte différentes caractéristiques comportementales des animaux. Pour les porcs et les rats, par exemple, l’attrait de ces animaux pour la nouveauté était l’élément clé. Chez ces campagnols, l’attrait pour des femelles en période d’activité sexuelle a été utilisé comme objet d’étude. Ainsi, la mémoire épisodique n’est pas liée à un sens ou à un comportement, mais à la capacité fondamentale intrinsèque de nombreuses espèces d’animaux de mémoriser des évènements spécifiques.
Plusieurs auteurs ont fait remarquer que la mémoire épisodique est importante pour une planification du futur (Raby et Clayton, 2009 ; Raby et al., 2007 ; Roberts, 2007 ; Crystal, 2010). Le fait d’être apte à détecter des règles dans l’occurrence d’évènements passés permet d’utiliser ces compétences pour « prédire » l’avenir. Depuis longtemps, des pics et des écureuils ont été observés cachant des fruits pour les consommer en période de pénurie. Il avait été dit cependant que ce comportement pourrait ne traduire que l’aptitude à cacher des aliments qu’ils auraient pu consommer immédiatement. Il n’était donc pas possible de conclure qu’ils le faisaient pour préparer l’avenir. Ce contre-argument a été levé par des expérimentations complémentaires.
Dans la continuité des expérimentations sur la mémoire épisodique chez les geais buissonniers, une nouvelle expérimentation conclut à la possibilité pour ces oiseaux de planifier leur futur. Dans celle-ci les geais buissonniers avaient à leur disposition deux types d’aliments, des arachides et des croquettes. Ils cherchent toujours à consommer un mélange des deux aliments. Une journée donnée, chaque oiseau a été placé dans une cage comprenant deux compartiments. Il a pu cacher ces aliments dans les deux compartiments. Le soir, l’animal a été maintenu dans un des compartiments et le lendemain matin il n’a reçu qu’un seul type d’aliment. Le jour suivant il a pu de nouveau cacher les deux aliments dans les deux compartiments. Il a été de nouveau confiné dans le même compartiment que la veille. On observe alors qu’il a caché préférentiellement l’aliment auquel il n’avait pas eu accès la veille dans le compartiment où il avait été confiné (Raby et al., 2007). Une autre expérimentation conduite sur un mustélidé sud-américain, le tayra (martre à tête grise, Eira barbara), a montré une aptitude à planifier des actions futures (Soley et Alvarado-Diaz, 2011). Ces animaux sont friands de bananes plantain mûres. Ils cachent ces fruits lorsqu’ils ne sont pas comestibles et ne les récupèrent et ne les consomment que lorsqu’ils sont devenus comestibles.
Une autre aptitude a été présentée comme étant spécifique de la conscience. Il s’agit de l’aptitude à connaître ses propres connaissances, ou métacognition. Les humains peuvent dire « je sais que je connais cela » ou « je l’ai sur le bout de la langue ». Tout le monde a en tête la description du souvenir de la consommation d’une madeleine par Proust (1913). Ce type de cheminement est difficile à tracer chez les animaux, en l’absence de langage. Des dispositifs spécifiques ont néanmoins permis de caractériser cette compétence chez certaines espèces. Par exemple, des animaux apprennent à appuyer sur des boutons. Deux représentations différentes, comme des mélanges d’images, d’odeurs ou de sons, sont proposées à des sujets animaux, notamment des dauphins (Smith et al., 1995). Si le sujet choisit une représentation il obtient ainsi une récompense, et il n’obtient rien s’il choisit l’autre image. Au début des essais, les deux représentations sont très différentes. Par la suite, elles sont de plus en plus semblables. Un troisième bouton est proposé aux sujets. L’appui sur ce dernier bouton permet d’obtenir une petite récompense dans tous les cas, quelle que soit leur réponse. Dans ces conditions, les sujets appuient plus sur le troisième bouton lorsque les deux représentations deviennent difficiles à discriminer. Ils montrent ainsi qu’ils peuvent mobiliser leurs connaissances.
Les dauphins, notamment, devaient reconnaître des sons de fréquences différentes en appuyant sur deux boutons spécifiques et avaient à disposition un bouton « joker » (Smith et al., 1995). Ils ont appuyé sur ce troisième bouton lorsqu’ils ont eu des difficultés à différencier les fréquences. La même conclusion est tirée d’études qui portaient sur des singes (Morgan et al., 2014), des rats (Foote et Cristal, 2007) et même des abeilles (Perry et Barron, 2013). Le dispositif a pu être complexifié, par exemple en posant la question aux animaux a posteriori après qu’ils aient fait leur choix. De plus, Hampton (2019) conclut que les macaques peuvent éviter de répondre à des tests de mémorisation quand ils les ont oubliés, chercher des informations supplémentaires quand elles leur manquent, et faire des paris cohérents après avoir fait des jugements.
Enfin, parmi les nombreux résultats portant sur la conscience des animaux, les relations interindividuelles occupent une place particulière. On sait que les animaux doivent lors de ces interactions constamment ajuster leurs actions en fonction de celles des partenaires. Des dispositifs spécifiques ont permis de tester des hypothèses liées à la réflexivité de l’individu, non seulement sur ses propres connaissances mais également sur celles de ses congénères. Il a été montré chez certaines espèces sociales qu’il existe une reconnaissance individuelle, par exemple sur le mode acoustique (Lengagne et al., 1999) et sur le mode visuel (Tibbets, 2002 ; Dasser, 1987 ; Coulon et al., 2009 ; Autier-Derian et al., 2013). Dans le contexte dyadique où il existe des relations de dominance et de subordination, un des deux individus a accès prioritairement à des ressources, alimentaires, environnementales ou sexuelles. On observe souvent une quasi-transitivité des relations de dominance et de subordination. Si un individu A domine un individu B et que B domine C, et si A domine également C, on parle de transitivité. Cependant, des expérimentations ont été conduites sur un groupe de macaques japonais (Macaca fuscata) qui était organisé en lignées maternelles. Ces expérimentations ont montré l’importance des alliances entre individus dans le cadre de ces relations de dominance et de subordination : deux individus de la lignée la plus subordonnée pouvaient dominer un individu seul d’une lignée plus dominante (Chapais, 1988). De plus, des observations ont été faites sur des singes capucins (Perry et al., 2004) et des hyènes (Engh et al., 2005). Elles ont montré que, dans ces deux espèces, deux subordonnés peuvent s’allier pour remporter les ressources contre un individu qui leur est dominant. Les résultats de ces observations montrent que les subordonnés s’allient non seulement en se connaissant mais en connaissant leurs statuts respectifs.
D’autres observations montrent que les corneilles (Bugnyar et Heinrich, 2005) et les geais buissonniers (Dally et al., 2005) modifient leurs stratégies de cache selon les observateurs. Call et Tomasello (2008) ont montré expérimentalement que des chimpanzés comprennent à la fois les objectifs et les intentions d’autres individus, y compris humains. D’autres textes rapportent l’aptitude de certains animaux, dont des oiseaux (corvidés et gallinacés), des singes et des rongeurs à « mentir ». Ainsi, des expériences chez les gallinacés ont mis en évidence un effet « d’audience » (Gyger et al., 1986 ; figure 3.3). Le coq n’a émis un cri d’alarme que s’il était en présence de poules. Il est resté silencieux si l’autre partie de la cage était vide ou contenait une caille. Marler et ses collègues (1986) rapportent le comportement de coqs placés dans un petit enclos qui était entouré soit de poules, soit de coqs. Lorsque ce sont des poules, le sujet émettait un cri d’alarme et allait se mettre à l’abri. Lorsqu’il était en présence de coqs, ce même sujet s’est mis à l’abri sans émettre de cri. Ces auteurs qualifient ce comportement de mensonge.
D’autres situations ont été explorées qui révèlent les capacités en termes de conscience de certaines espèces animales. Ainsi, des primates non humains (Boesch et Boesch, 1983 ; Whiten et al., 1999 ; Fragaszy et al., 2004 ; Visalberghi et al., 2009), mais aussi des oiseaux comme le corbeau de Nouvelle Calédonie ou Corvus moneduloides (von Bayern et al., 2018) ont montré leurs capacités à utiliser des outils pour accéder à des aliments.
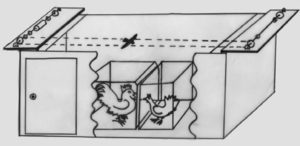
Dispositif expérimental : la cage où est enfermé le coq est placée dans une grande pièce. Un dispositif permet de faire passer au-dessus de la cage une silhouette de rapace. Une autre cage est placée en face de la cage du coq. Dans cette seconde cage, est placée en fonction des tests, soit une poule, soit une caille, soit cette cage est laissée vide. Dans d’autres expériences, plusieurs poules ou des coqs sont mis en présence du coq-sujet.
Source : Vivre parmi les animaux, mieux les comprendre de Pierre Le Neindre et Bertrand L. Deputte, paru aux éditions Quæ




